当我们仰望星空时,我们常常惊叹于群星的繁丽与宇宙的广袤,惊叹于在那深邃无边的空间里,究竟隐藏着多少人类未知的秘密。然而,就在我们每一个人的身体里,也隐藏着一个同样复杂且神秘的宇宙:大脑。我们的大脑包含大约一千亿(1011)个神经元——这个数字是整个银河系恒星数目的10倍——而其中平均每个神经元又与其他神经元形成约一千个被称为“突触”的联接节点。正是通过这些突触,一个神经元得以接受来自其他神经元的输入信息(兴奋性的或抑制性的),将其整合、处理,产生自身的输出信息,并通过轴突将其传递给下一级的神经元。也正是通过这些突触,这一千亿个神经元彼此之间相互联接,形成数以万计的神经环路,进而构成一个复杂且庞大的神经网络系统。在这个网络中的神经环路上,编码着各种信息的神经冲动一刻不停地奔腾着。当我们的眼、耳、鼻、手接触到外部世界的影像、声音、气味、材质,这些感觉刺激通过我们的感官转换为神经环路上的电化学信号,一路奔涌进入我们的大脑,让我们具有了视觉、听觉、嗅觉和触觉。而当我们需要对这些外界刺激做出响应,带有“指令”信息的神经信号又从中枢飞奔至身体的各部位,驱使我们的身体产生各种自觉或不自觉的反应。
然而,生物体的大脑与外部世界的“互动”远远不止上述的简单信息交换。科学家早已发现,大脑的某些功能会在外界刺激的影响下发生改变,即大脑具有可塑性。例如,普通人经过训练,逐渐能够分辨出原来不能分辨的色彩、形状、味道或质地。同样,人也能够通过学习获得一些永久的技能,比如说某种语言、骑自行车或者弹钢琴。对这些技能的学习往往具有一定的“关键期”,错过了这个关键期之后,学习的效果会显著下降,甚至是根本学不会。大脑的这些可塑的特性提示,脑中神经元的相互联接状态——也就是那些负责感觉、运动、记忆、情绪的神经环路——并非一成不变,而是不断地在来自外界刺激的“调教”下进行修正、重塑。正如人们所能观察到,即便是在基因水平上完全相同的同卵双胞胎,也会发展出拥有不同能力、个性和情感的独立个体,提示他们脑中神经环路的联接状态是不同的。换而言之,正是来自外界的不同经历和生活体验把我们塑造成了一个个独特的个体。然而,这种塑造作用是如何发生的,或者用神经科学的术语描述,外界刺激如何影响神经环路的形成与可塑性,一直是神经科学家苦苦探寻但却尚未完全解答的问题。
前面提到,神经元之间的信息传递通过突触实现,因此突触也可以被看作是神经环路的基本节点。大脑中绝大部分的兴奋性突触位于神经元的树突上,并且由两部分构成:负责传出信息的是来自上一级神经元的轴突末端,而负责接收信息的是下一级神经元树突上一种“棘刺”状的凸出结构,被称作“树突棘”。这些树突棘最初是由被称为“现代神经科学之父”的十九世纪西班牙神经解剖学家Santiago Ramón y Cajal用手绘的方式记录与描述(图1)。而树突棘做为突触后的主要结构进行神经信息传递的功能在二十世纪中晚期才通过电镜、电生理、光学成像等方法得以验证。通过分析神经元上树突棘的数量与形状,科学家可以估算出神经环路中联接的多少与强弱。研究发现,树突棘的数量在出生后早期急速增加,提示在幼年期的发育过程中神经元之间形成大量的突触联接。有趣的是在个体经由青春期逐渐进入成年期的过程中,树突棘的总数量反而显著减少,也就是说已形成的联接会被“修剪”,使整个神经网络的联接更加精确(图2)。这一“树突棘修剪”过程对大脑的正常功能至关重要,在例如孤独症、精神分裂症等发育性神经系统疾病中均发现了树突棘修剪的异常。然而,关于什么分子机制调控了这一修剪过程,并在修剪过程中决定了树突棘的不同命运——被修剪掉或者得以存活——我们依然所知甚少。这些正是中国科学院神经科学研究所于翔研究组感兴趣的科学问题。
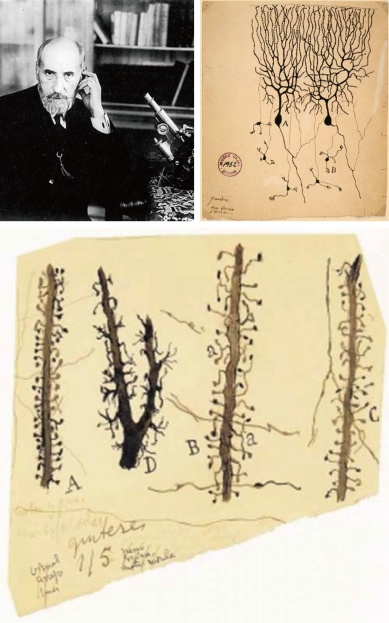
图1:西班牙著名的神经生物学家Santiago Ramóny Cajal(1852-1934)与其手绘的小脑浦肯野神经元以及不同形态的树突棘。图片引自参考文献[1]。
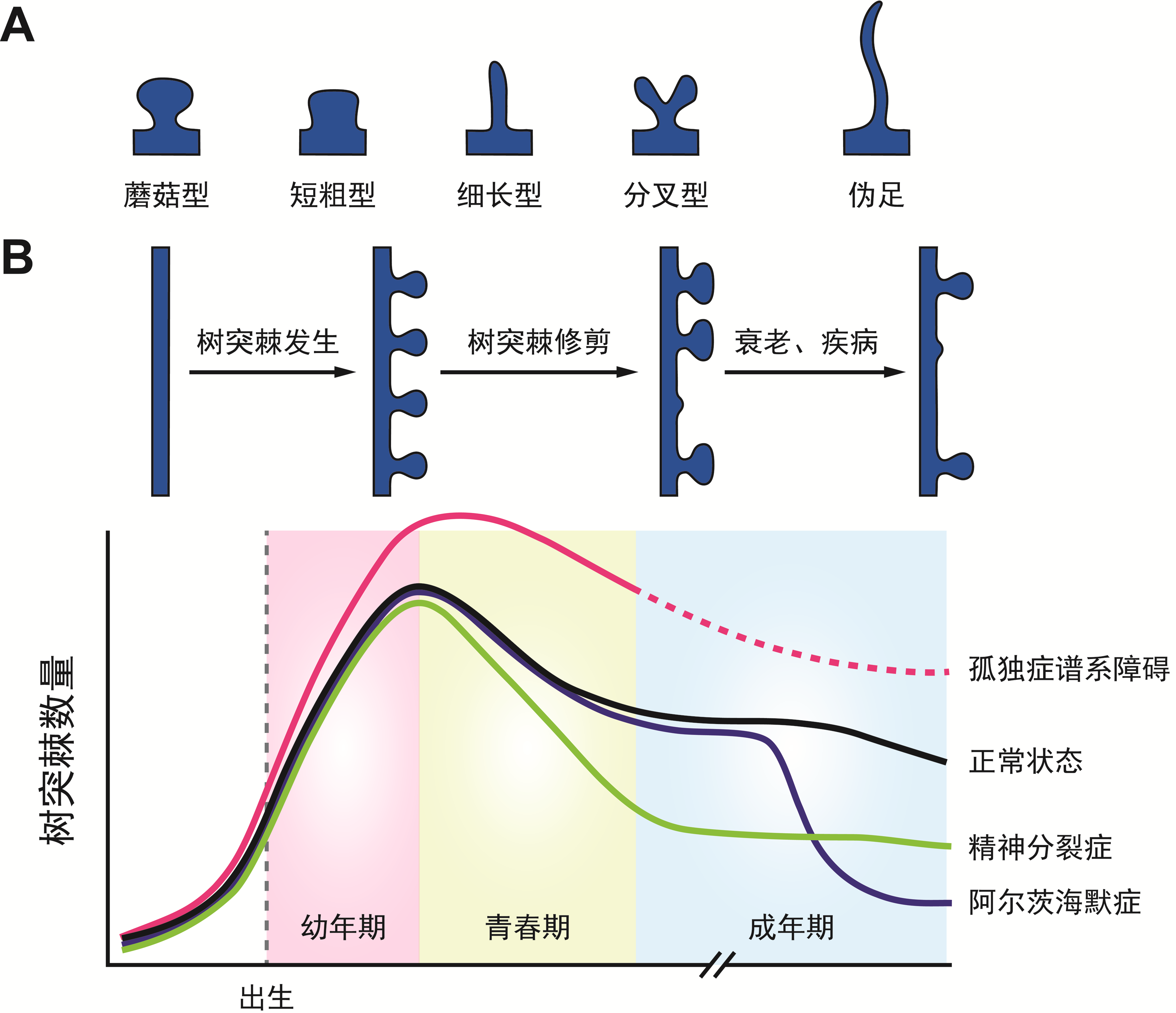
图2:A:四种树突棘类型和伪足的形态示意图。B:正常发育过程中(黑线)以及孤独症(粉色)、精神分裂症(绿色)、阿尔茨海默症(蓝色)等疾病状态下,脑内树突棘数量的变化。图片引自参考文献[1]。
小鼠的触须是一种非常发达的感觉器官。小鼠的觅食、探索、感知危险以及其他日常活动都有赖于其触须的感觉输入。因此,于翔研究组利用与小鼠触须感觉对应的大脑感觉区域——桶状皮层——作为模型尝试解答上述问题。他们首先发现该脑区在发育过程中的树突棘修剪受触须感觉经验的双向调控,并且与树突棘形态的成熟度协同变化——通过丰富环境饲养增加小鼠的感觉输入能够同步加速树突棘的修剪与其形态的成熟,而剪除触须则同时阻止这两个过程的进行。他们据此推测树突棘的修剪与成熟是由同一种竞争机制所介导:相邻的树突棘会竞争胞内的某种分子资源,获胜的树突棘得以存留下来并走向成熟,而失败的一方则接受被修剪的惩罚。
那么这种分子资源到底是什么?在探寻这种分子资源的真实身份的过程中,一类独特的分子,cadherin/catenin细胞粘附复合物(简称3C复合物),跃入眼帘。这是一类位于突触的递质释放区域与受体表达区域的蛋白复合物,由三个主要成员相互结合而成:N-cadherin、β-catenin和αN-catenin。N-cadherin是一种跨膜蛋白,树突棘上N-cadherin的胞外段会伸到突触间隙中和轴突末端的N-cadherin胞外段“搭”在一起,如同魔力粘扣一样形成粘附的效果。而在树突棘细胞膜的另一面,N-cadherin的胞内段和β-catenin、αN-catenin等蛋白组分组成蛋白复合体,招募肌动蛋白骨架聚集,从而为树突棘的结构提供了物理性的支撑。尤为重要的是,3C复合物能够响应神经电活动发生上膜、内吞等变化,从而将突触前电活动输入的变化和突触后树突棘内细胞骨架的变化相互关联。
于翔研究组通过对体外培养的神经元进行活细胞成像,发现在单个树突棘上局部富集3C复合物可以同时引起被操作树突棘的增大和相邻树突棘的缩小或消失(图3)。通过光遗传学手段激活单个树突棘并与相邻未激活的树突棘进行比较,发现了类似的树突棘命运分化:前者增大而后者被修剪,并且这一过程依赖于3C复合物在两个树突棘中的重新分布。这种树突棘的命运分化取决于两个树突棘之间的物理距离和N-cadherin(3C复合物的重要组分)的运动性,而不依赖于蛋白合成或降解,进一步说明两个树突棘是对现存的3C复合物进行竞争,而不是各自对其进行合成或降解。
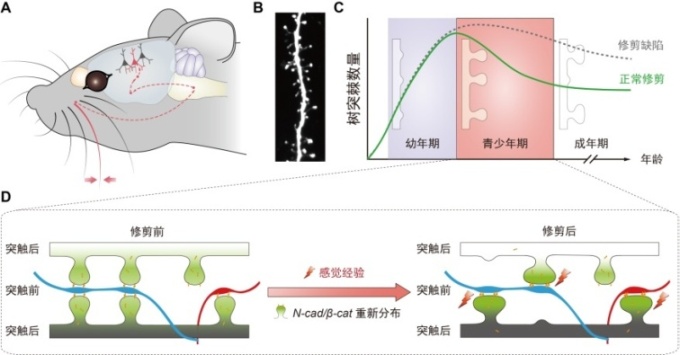
图3:(A) 小鼠触须接受到的感觉刺激传入到躯体感觉皮层神经元(虚线为其传导途径)。(B)躯体感觉皮层桶状区域第2/3层锥体神经元的树突棘示例。(C) 躯体感觉皮层神经元树突棘数目在正常(绿色)和修剪缺陷(灰色)情况下随年龄变化的趋势。(D) Cadherin/catenin复合物介导树突棘修剪的竞争模型:由感觉经验所带来的神经电活动增强导致相邻树突棘之间相互竞争胞内的cadherin/catenin复合物,这一竞争导致后者在树突棘之间的重新分布:获得cadherin/catenin复合物较多的树突棘得以存留、成熟,而失去该复合物的树突棘则被修剪。
在体条件下,通过结合小鼠基因操作和定点病毒注射,于翔研究组使小鼠桶状皮层中的一部分树突棘在突触前拥有更高的3C粘附水平。他们发现在经历了正常修剪之后,这些树突棘更多的被保留下来,并且形态更为成熟,而其邻近的低粘附的树突棘则更多地被修剪掉。进一步的实验表明感觉经验对于树突棘修剪和成熟的调控也依赖于3C复合物的作用。
虽然我们离完全解开大脑可塑性的秘密还很遥远,但是中科院神经所于翔研究组的这项研究使我们在这条路上前进了一小步。通过这项工作,我们知道,在青春期的神经环路精确化过程中,有一类关键且有限的分子资源。这类能够促进细胞粘附的3C复合物能够加强树突棘和它的突触前“小伙伴”——轴突末端——之间的粘附作用,从而使树突棘变得更加稳定。然而好东西总是稀少的,细胞内正常的3C复合物并不足以满足每一个树突棘的需求,因此相邻的树突棘之间就会产生对于这类分子的竞争。那些接受强输入的树突棘更容易在这样的竞争中胜出,抢夺到更多的3C粘附复合物。一旦最初的优势形成便一发而不可收拾,越来越多的3C复合物以一种正反馈的方式不断积累到获胜的树突棘中,而那个失败的树突棘不断失去3C复合物,只能任由自己的结构松散、崩溃,最后难逃被修剪的命运。神经系统以这样一种简单的竞争机制保证了使用频率较高、输入较强的环路联接被保留下来并加以强化,而使用频率低、输入较弱的联接被去除,从而使系统资源得到最优化的分配,神经环路的联接更加精确。这一基于有限分子资源的竞争分配机制在亚细胞的层面印证了那条“用之或弃之”的神经科学基本概念,拓展了我们对于神经环路精确化和大脑可塑性的理解,并且很可能代表了生物系统的一种普遍策略。
[1]边文杰,于翔. 树突棘的动态变化与调控机制. 生命科学, 2015, 27(3): 294-305
(审核:顾勇、何杰)
上一篇: Nature子刊:不一样的神经元修剪
下一篇: 西方54个学习理论你知道多少个?